Striving, Playing and Learning
On aggression and the dynamics of learning in the play and struggle called 'life'
By Popko P. van der Molen & Johan M.G. van der Dennen
Presented at the 1978 Congress of the "Ethologische Gesellschaft" at Basel and at the First Congress of the European Section of the International Society for the Research on Aggression (I.S.R.A.) at Strassburg, sept. 1981.
*** This page is under construction *** (figuren nog toevoegen)
Contents
- 1 Table of Contents:
- 2 Summary
- 3 1. Critical recovery periods, an Experiment
- 4 1.1 Hormonal Feedback
- 5 1.2 Recovery Intervals and Learning
- 6 2. Some models of agonistic behaviour
- 7 2.1 The Conflict- or Ambivalence Hypothesis
- 8 2.2 The Psychohydraulic Model: the Catharsis Hypothesis
- 9 2.3 Aggression and Emergency Behavior
- 10 2.4 Aggression,and Behavioral Reduction versus Behavioral Expansion
- 11 2.5 Butterfly Catastrophes
- 12 3. The Reversal Hypothesis
- 13 4. An Additional Hypothesis: Integration of Theories on Aggression and on Learning
- 14 5. Positive and Negative Learning Spirals
- 15 5.1 Genetical Aspects and 'Framing'
- 16 5.2 Growth of Skills versus Growth of Unskills
- 17 5.3 Reversability of 'Unskills'
- 18 5.4 Displacement and Neuroses versus Psychological Health
- 19 6. The Use of the 'Aggression'-Label as an Element of the Agonistic Repertoire
- 20 7. By way of Conclusion
- 21 References
Table of Contents:
Summary
1. Critical recovery periods, an Experiment
1.1 Hormonal Feedback
1.2 Recovery Intervals and Learning
2. Some models of agonistic behaviour
2.1 The Conflict- or Ambivalence Hypothesis
2.2 The Psychohydraulic Model: the Catharsis Hypothesis
2.3 Aggression and Emergency Behavior
2.4 Aggression,and Behavioral Reduction versus Behavioral Expansion
3. The Reversal Hypothesis
4. An Additional Hypothesis: Integration of Theories on Aggression and on Learning
5. Positive and Negative Learning Spirals
6. The Use of the 'Aggression'-Label as an Element of the Agonistic Repertoire
7. By way of Conclusion
Summary
✰✰ <level 2> A novel model on the dynamics of learning is presented which bears particular significance to the notion of (violent) aggressiveness. It suggests, among other things, that aggressive behavior cannot be properly understood whithout considering the basic dynamics of learning involved, without recognizing the shifting balance between 'striving and playing' and without recognizing the prevailing balance between social skills and 'unskills'. If any of these aspects are not considered, 'Aggression' or 'aggressiveness' will tend to be viewed as a separate phenomenon in its own right - which is unfortunately a widespread habit, not only in daily life, but also among the very investigators of the subject. The very use of labels such as 'Aggressiveness' and 'violence' keeps our attention trapped and hampers awareness of the underlying dynamic behavioral processes . In the first half of this paper some well-known models of aggression are critically reviewed and some of the basic concepts are subsequently integrated with theories of learning and with the 'reversal' hypothesis of antagonistic motivations, which replaces the optimum-arousal theory. An additional hypothesis on the long-term regulation of attention then completes the emerging model of learning. A model which stresses the antagonism between the possible growth of stereotyped and inflexible emergency reactions, and the long-term growth of varieties of subtle and flexible coping-response patterns (skills). The proposed model conflicts with many classical learning-conditioning concepts in that it predicts that the prevailing emotional (telic/paratelic, or centripetal/centrifugal) balance determines whether certain aversive and/or ambivalent stimuli result in the long run in attraction or in repulsion.
1. Critical recovery periods, an Experiment
✰✰✰ <level 3> In a series of experiments (Van der Molen, 1979, 198]), approximately 60 male housemice of the CPB's strain were weaned and isolated at the age of 21 days. Subsequently they were introduced every day or every other day into an unfamiliar observation cage, populated by a group of resident mice showing territorial aggressive behavior towards strangers. Being outnumbered by at least four to one and being on foreign territory, a trainee never had a chance to win and therefore was beaten up every time when introduced, and often slightly wounded. After 5 to 10 days of training, these trainees would immediately show avoidance behavior when being introduced, even when not under attack by any resident mouse at all.
There seemed, however, to be some sort of critical training frequency, i.e., if the periods of rest in isolation between the training introductions were too long, the effect of the whole training was the opposite of what was intended in the first place. If an 'experienced animal' or an experienced 'fleeing' trainee was introduced once every week or once every other week instead of every day, he would tend to put up skilled resistance. Such a male did not attack residents, but refrained from 'fleeing frantically' and appeared to 'stand firm' when necessary. By manipulating only the 'training frequency', trainees could be transformed from inexperienced males into either 'fearful fleeing males' or 'skilled fighters'. It should be remembered that neither type of trainee ever won a fight. During every introduction in their training period they had to fight against heavy odds. 'Skilled fighters' simply managed to hold out and fight back now and then without 'running for their lives' frantically. With the procedure described above trainees became either more skilled in fighting or more fearful than inexperienced males. Furthermore, in-experienced males varied more in their reaction patterns than did either of the two classes of trainees. Once a male had become a 'skilled fighter' it was very difficult to reshape him into a 'fearful fleeer' again. Apparently the same type of experience can lead to different learning processes, depending on the input frequency of the experience. The one direction in the learning spiral (deviation-amplifying mutual causal process: Maruyama, 1963) leads to stereotyped and intensive avoidance reactions; the other direction leads to increased skill in offering resistance.
Thus the temporal pattern of experience appears to be of great importance for its ultimate effect. Besides, individual mice differed in their critical digestion time for this type of experience; for the CPB's strain males in question it varied between 2 and 6 days. In general, males with a background of 'normal' social experience were rather well able to hold out against avalanches of territorial aggression. A 'normal' social history apparently provided ample opportunity for these mice to 'digest' their aggressive encounters in such a way that they became skilled in dealing adequately with agonistic social situations.
The following conclusions were drawn from these experiments:
(1) Male housemice can easily be trained in such a way that they perform a uniform type of avoidance behavior when introduced into a strange environment, inhabited by resident mice.
(2) The training required for turning an inexperienced male into a 'coward' is almost the same as the training needed to turn him into a 'hero'. The only difference is that in the former training schedule less 'digestion time' is available for every experience than in the latter.
(3) This 'critical digestion time' varies somewhat between individuals, and depends i.a. on their life-history.
1.1 Hormonal Feedback
✰✰✰ <level 3> A possible explanation of (part of) the phenomenon of 'critical frequencies of experience' may be found in the influence of fearful experiences on the endocrine system. Fear reduces for instance the testosterone level and with that probably the tendency to put up resistance when provoked (Bermond, 1977). Long training intervals allow this and other hormone levels to return to normal, but when the intervals between the fearful experiences are too short, the effects accumulate, thus rendering the individual inclined to flee instead of putting up resistance when challenged again. A considerable amount of experimental evidence is available to support this idea. For instance, Eleftheriou & Church (1968) demonstrated that in mice the blood plasma levels of LH, LH-RF and FSH decreased after aggressive encounters. The hormonal changes in losers appeared to be cumulative in contrast to the hormonal changes in winner-mice. Rose et al. (1972) demonstrated similar hormonal effects in rhesus monkeys. Many investigators (e.g. Bronson et al., 1973; Leshner, 1975; Schuurman, 1981, and in: Wiepkema & Van Hooff, 1977) found that aggressive encounters brought about stronger physiological changes (i.a., in blood sugar level) in losers than in winners. In more anthropomorphic terms: A conflict or encounter which elicits emergency reactions and raises anxiety in the subordinate may well represent an ordinary - eventually exciting - matter of routine to the dominant. All this means that apart from opposite conditioning effects of aggressive encounters on winners and losers on the neural level, a stable behavioral divergence between the social roles is still further enhanced and consolidated by different hormonal adaptations (Leshner, 1975). Similar effects have also been demonstrated in humans. Kreuz et al. (1972) conducted physio¬logical studies on trainees in a military camp and found that in periods of high tension, stress and extremely hard work, their testosterone level decreased considerably and recovered only gradually after relaxation.
1.2 Recovery Intervals and Learning
✰✰✰ <level 3> Apart from the hormonal feedback as depicted above, differential 'learning' processes on the neural level are involved as well (Leshner, 1975). Because animals in a Bèta-role and animals in an Alpha-role experience aggressive encounters in a different way (losing and winning resp.), different neural information is stored and processed. Besides, as e.g. Buchholtz (1978) and Trincker (1966) point out, there is a great functional and physiological difference between short-term memory storage and long-term memory storage. And there are limits to the amount of experiences of a certain type pro time unit that an individual can sucessfully digest and transform into (greater) long-term skills. This phenomenon has been established in a wide range of species from insects (Gross, 1978) to Man (Buss, 1973, pp. 278-280). The neural and hormonal feedback systems function synergistically. Whereas in lower developed species hormonal changes are relatively important, we may expect that in neurally higher evolved species neural mechanisms of adaptation will be relatively important in comparison with the purely hormonal mechanisms.
2. Some models of agonistic behaviour
✰✰✰ <level 3> Below, a general model is construed that covers adaptational mechanisms of various complexities and various functional levels, hormonal as well as neural, and including the higher learning processes in Man. In order to arrive at that model, we first proceed with reviewing some current models on aggression and on agonistic behaviour in general. For it is assumed that in particular those learning processes are of paramount importance for the individual, that lead to skills in dealing with agonistic encounters, other aspects of role-conflict, etc.
2.1 The Conflict- or Ambivalence Hypothesis
✰✰✰ <level 3> Somewhat oversimplified, this hypothesis states that agonistic and related behaviors are the product of activation of, and interactions between, two major antagonistic 'drives1 or 'tendencies': the attack- or approach-tendency and escape- or withdrawal-tendency. Many courting behaviors may similarly be considered as the product of activation and interactions of 3 tendencies, viz. the attack-, escape- and sex-tendency (see e.g., Tinbergen, 1952; Baerends et al., 1955; Kruijt, 1964; Hinde, 1966; Baerends, 1975). 'Approach' and 'withdrawal' behaviors may shade off into one another through intermediate - compromise - types of response, depending on the balance between the attack- and escape-tendency. Such compromise movements or postures are often postulated to be the evolutionary origin of a communicative signal or display. If the basic tendencies are highly activated, the antagonist behavior types tend to change in a more sudden and dramatic way than if they are moderately activated. In the former case attack may suddenly change into escape and escape may suddenly (catastrophically) change into attack.
This state of affairs may graphically be presented and mathematically described with the aid of the bistable models from the mathemathical branch of catastrophe theory as presented by e.g. Thorn & Zeeman (1974) and Zeeman (1976) (see Fig. 1). This model of agonistic behavior, however, accounts only for short-term changes in mood and behavior. No predictions are implied that reach beyond the particular agonistic setting under scope. The catharsis hypothesis, on the other hand, is primarily concerned with long-term predictions.

2.2 The Psychohydraulic Model: the Catharsis Hypothesis
✰✰✰ <level 3> The catharsis hypothesis, when applied to aggression (Lorenz, 1950; Leyhausen, 1967; Eibl-Eibesfeldt, 1975), states that the longer an individual has not shown aggressive behavior, the easier aggressive behavior is evoked by external stimuli. Eventually the aggression-deprived individual will even start to seek opportunities to direct his aggressive behavior towards something or other and thus release his internally stored 'aggressive energy', ultimately to the point of releasing it 'in vacuo'. This process can be visualized through what has been called the 'psychohydraulic model', or, less respectfully, the toilet flush model.
Whereas this model may be quite adequate for describing some other functions like for example male sexual tendencies or feeding behavior, its use for describing aggressive behavior has become somewhat obsolete (Hinde, 1960, 1966; Manning, 1969; van Dijk, 1977; Zillman, 1979, van der Dennen, 1980; van der Molen, 1981); mainly because little evidence has been found that long-term deprivation of an opportunity to fight causes an increase in the readiness with which fighting can specifically be elicted. Yet appetitive behavior for agonistic interaction can nonetheless be demonstrated in a wide range of species: fish (Thompson, 1963; Rasa, 1971; Sevenster, 1973), birds (Thompson, 1964; Cole & Parker, 1971; Cherek et al., 1973), mice (Lagerspetz, 1964; Tellegen et al., 1969; Legrand, 1970; Tellegen & Horn, 1972; Connor, 1974; Kelsey & Cassidy, 1976; Connor & Watson, 1977), rats (Dreyer & Church, 1970; Taylor, 1975), hamsters (Eibl-Eibesfeldt, 1971), and monkeys (Azrin et al., 1965).
But in every case investigated, such alleged appetence for aggressive behavior could equally well be labeled as appetence for certain other functional pattern such as e.g. territorial behavior and/or as attempts of the individual to maintain an optimum level of arousal. "Excitement and kinetic activity are shown to be dependent on the external stimulus state of the animals decreasing under conditions of low mean environmental stimulation. These findings indicate that an animal attempts to regulate its internal stimulus state by behavioural means when the component variables of this state have been disturbed by environmental conditions" (Rasa, 1971).
Sevenster (pers. comm., 1980) arrived at similar conclusions in his experiments with sticklebacks, when evaluating his results on the rewarding effect of opportunities to interact aggressively. He investigated the rewarding properties of each component of the situation of agonistic interaction in question separately, and concluded that it is, in particular, the element of environmental change which works rewarding for the (somewhat arousal-deprived) male stickleback, but only so, if the change in question induces no flight behavior. Parallel to these findings are experimental results as those of Kavanau (1967), who showed poignantly how mice, being forced to respond in stereotyped experimental situations, will give 'incorrect' responses as a means of introducing variability.
From all these considerations we may conclude that for understanding function and (long-term) causation of aggressive behavior it is apparently of great importance to include phenomena such as 'boredom', 'excitement- seeking' and 'anxiety' in our models.
Specific appetence for aggressive behavior may in fact not exist at all in ordinary natural settings in which plenty of opportunity is available for attaining proper arousal levels. This is also suggested by, for instance, Goodall's (1971) descriptions of the occurrence of aggressive encounters in the daily life of free living chimpanzees. The frequency of intraspecific aggressive behavior varies strongly for every individual and for the whole group in question, and is highest whenever hierarchical relations have become unclear. The readiness to show intraspecific aggres¬sive behavior does not seem to depend at all on the period of time that has elapsed since the last agonistic confrontation, but rather on circumstances which make it difficult to avoid the application of coercive means.
The existence of a drive for aggression, irrespective of a functional context, would, in fact, be highly improbable from an evolutionary point of view (Craig, 1918, 1928; Tinbergen, 1956; Hinde, 1960, 1970, 1974; Marler & Hamilton, 1966; Wilson, 1971, 1975; Scott, 1973; Crook, 1973; Schuster, 1978), the rationale of which has been analyzed in game-theoretical and 'Evolutionary Stable Strategy' terms (Hamilton, 1971; Maynard- Smith, 1974, 1978; Maynard-Smith & Price, 1973; Parker, 1974; see also: Wilson, 1975; Dawkins, 1976; Fry, 1980). An individual runs heavy risks every time he engages in aggressive encounters. Therefore it seems good strategy to reserve aggressive behavior for situations in which the risk of damage is sufficiently counterbalanced by a possible raise in the chances of survival and/or propagation of the specific genetic information in question, after successful agonistic action.
It would appear that the regularity with which the males of certain species carry out charging displays can be more adequately explained by the 'hierarchico-cybernetic model' with the aggression-system as a low- level subroutine (Baerends, I960; Tinbergen, 1969; van Dijk, 1977) than by an energy model of aggressive motivation (Lorenz, 1963; Rasa, 1971, 1980).
2.3 Aggression and Emergency Behavior
✰✰✰ <level 3> Van Rooijen (1976) reviewed models of 'escape-' and 'emergency-' reactions as derived from current ethological and psychological research. In his study he described a general 'emergency-system' which prepares individuals for responding adequately in cases of emergency, and he labels it 'Anticipation of Damage System' (ADS). An essential of van Rooijen's model is the application of a strategy of 'flexibel response'. The more an individual is threatened, the stronger his ADS is activated. The level of activation determines which types of behavior can be expected. Furthermore, individuals can 'learn' which situations are best met with a careful approach and which situations require instantaneous flight or attack. Van Rooijen suggests that the learning processes themselves are probably enhanced most by a moderate activation level of the emergency system. Considerable information uptake- and processing capacity is required for learning, whereas in the most extreme emergency-reaction-modes such as e.g. panicky flight, the full neural processing capacity is probably reserved for ad hoc problem-solving. When the ADS is activated too strongly and/or too continuously, pathogenetic effects emerge, such as neuroses, phobias, psychosomatoses, and even physical disorders (Blythe, 1973). These pathological traits in their turn enhance, through inadequately rigid responses in an ever-varying environment, strong stimulation of the emergency system. Thus vicious spirals ('Teufelskreize') of ever increasing pathogenic effects are likely to occur as long as an individual does not often enough and long enough succeed in reducing the alarming stimuli and the resulting level of arousal.
Agonistic behaviors may be fitted into van Rooijen's model in two ways. On the one hand, flight and attack may be final - suddenly and catastrophically occurring - reflexes during high activation of the emergency system. On the other hand, flight and, in particular, attack can be learned and may in fact be optional responses to specific familiar emergency situations. In that case flight and attack may already occur at a moderate activation level of the emergency system, and consequently tend to change more gradually into each other.
2.4 Aggression,and Behavioral Reduction versus Behavioral Expansion
✰✰✰ <level 3> Kortmulder's (1974) hypothesis of 'Behavioral Expansion' runs to some extent parallel to van Rooijen's ideas. To the classical postulate of antagonistic attack- and escape-tendencies, Kortmulder adds the antagonism between 'centrifugal' and 'centripetal' tendencies. Full-scale attack and flight are both considered to be centripetal behaviors whereas courtship and, in particular, playful (agonistic) interactions are considered to be more centrifugal types of behavior. The terms centrifugal and centripetal refer to the tendency of expanding the (interactive) behavioral activities, respectively to the tendency of reducing and simplifying the (interactive) behavior situation. Strong activation of the 'emergency system' (van Rooijen) corresponds with 'centripetal tendencies' (Kortmulder), and a weak activation of the 'emergency system' corresponds with 'centrifugal tendencies' of behavioral expansion.
When exploring the environment or when playfully interacting with a partner, an individual is supposed to be in a process of 'behavioral expansion' (centrifugal behavioral forces; cf. also Orme-Johnson's, 1973, 'Stay-and-play response'). But when the situation becomes dangerous - for instance when a partner suddenly play-attacks too frighteningly hard - the 'Anticipation of Damage System' is activated and an increased centripetal tendency leads to reduction of the behavioral setting through flight or attack.
According to Kortmulder, individuals strive after being in a state of behavioral expansion as often as possible. Through that behavioral state their behavioral repertoire expands and their skill in dealing with the animate and inanimate environment increases through experience.
Parallel to Maslow's (1968) behavioral models of men and other primates, Kortmulder (1972, 1974) discerns between (a) animals with a behavioral repertoire which is fully actualized through sufficient periods of behavioral expansion (self-actualizers), and (b) animals who have stayed under the influence of centripetal behavioral forces too often and too continuously and who subsequently show a reduced, 'truncated', and highly stereotyped behavioral repertoire. "Whereas self-actualizing fishes typically swim around actively also outside reproductive periods, or feed, inferior fishes often stand still for long periods of time, even in the absence of the dominant male. If they move, locomotion is typically, wavering and slow. They often do not feed at all ........ The general behaviour of such inferior fishes is somewhat similar to the behaviour of a sick fish. This is so even in the absence of any visible trace of bodily damage. The above observations suggest that the switch from self-actualizing to inferior behaviour marks the onset of a profound physiological change which is easily reversible when short-lasting but which tends towards irreversibility when submission is prolonged" (Kortmulder, 1972).
2.5 Butterfly Catastrophes
✰✰✰ <level 3> Taking the models of Kortmulder and van Rooijen into account, we can explain not only the gradual and the sudden changes from fight to escape and vice versa, but also the gradual and sudden changes from play to full-scale agonistic interaction. Thus we may integrate in one model the two different modes of aggressive interaction, viz. the 'rough- and-tumble' and the 'reactive fighting' as may be discerned in many species (rhesus monkeys: Harlow, 1965; chimpanzees: Goodall, 1971; human children: Blurton-Jones, 1967; Hamburg, 1971).
Like the mechanisms as postulated by the Conflict- or Ambivalence Hypothesis (see fig. 1), the mechanisms as described above may also be represented graphically (and mathematically) by means of a model from catastrophe-theory. Butterfly catastrophes (Zeeman, 1976) describe tri- stable systems, for instance three such behavioral modes which can shade off into one another, but which may, under certain conditions, also turn into each other in a sudden and catastrophic way (see fig. 2).

The dotted area indicates three possible behavioural modes. Horizontal lines indicate bistability between attack and escape and vertical lines indicate bistability between play and other behaviour. Catastrophic changes in behaviour may occur in half of the motivational states that are indicated by the lines on the bottom projection, depending on the previous motivational state.
One of the parameters of the graph in fig. 2 is the so-called 'butterfly-factor' which determines the size of the pocket in the bifurcation-set. A bigger pocket represents a greater probability of playful and explorative behavior. The 'butterfly-factor' might therefore be used to indicate an individual's skill in dealing with ambivalent motivational situations without having to resort to full-scale attack or flight. In other words: by an increasing butterfly-factor we may graphically represent the long-term effect of adequate frequencies of behavioral expansion, namely the raise in skills which results from the centrifugal behavioral activities of play and exploration.
A raise in skills causes in its turn a lower necessity to resort to centripetal behavior and thus a lower .probability of purely agonistic behavior and consequently a further increased probability of playful and explorative behavior.
This positive feedback mechanism in learning represents the opposite of the vicious spirals of ever-increasing pathogenic effects mentioned above.
Of course, it is also possible to block the positive learning-spiral by denying the individual any reasonable amount of experience, for instance by isolation. Centrifugal moods cannot then simply result in drifting into relevant experiences through lack of environmental possibilities. Individuals which have been isolated during long and crucial periods of their lives, generally exhibit inadequate social reactions (notably, anomalous sexual and agonistic behavior) to a high degree (jungle fowl: Kruijt, 1964; mice: van der Molen, 1979; rats: Peijs, 1977; rhesus monkeys: Harlow, 1961, Nowak & Harlow, 1975; squirrel monkeys: Hopf, 1981). As Kortmulder (1974) points out, courtship may be compared to play in that courtship partners 'learn' to get behaviorally adjusted to, and synchronized with, each other and thus 'learn' to avoid full-scale agonistic behavior towards one another. In the course of the courting process centripetal tendencies gradually give way to centrifugal tendencies until mutual aggression has virtually disappeared and the animals have learned to 'know' each other in playful interaction. Therefore Zeeman's graphical representation might 'mutatis mutandis' be used equally well to describe the interactions between sexual-, approach-, withdrawal-, attack- and escape-tendencies in courtship and the learning processes involved.
3. The Reversal Hypothesis
✰✰✰ <level 3> Kortmulder's hypothesis of 'Behavioral Expansion' was derived from animal behavior. A somewhat akin model was derived from studies on human behavior by Smith & Apter (1975) and Apter & Smith (1979). Kortmulder's antagonistic 'centripetal' and 'centrifugal' tendencies can be found in Smith & Apter's model as 'telic' (goal-directed) and 'paratelic' (behavior-directed) tendencies. Apter & Smith (1979) explain: "we shall simply start by stating one of the fundamental postulates of the theory. This is that certain psychological processes, especially certain motivational and emotional processes, exhibit bistability rather than homeostasis (i,e,; unistability). Switching from one stable state to the other in a bistable system can be reffered to as a reversal (hence the name of the theory) and may be brought about by a number of different factors. For example, it is argued that at certain times the individual seeks high arousal which is then felt as pleasant when achieved ('excitement'); at other times he reverses to a state in which he seeks low arousal, at which time high arousal is felt as unpleasant ('anxiety'). In the former case low arousal is felt as unpleasant ('boredom') and in the latter case low arousal is felt as pleasant ('relaxation'). This is summarized in the two hypothetical curves drawn in the accompanying diagram (Fig. 3a), each curve representing the relationship between arousal and affective tone for each of the two stable states. These two states are labeled 'telic' and 'paratelic' respectively. This view of the relationship between felt arousal and affective tone contrasts sharply with that of optimal arousal theory which is a homeostatic theory, although the typical inverted U-curve (shown dotted in Fig. 3a) postulated by optimal arousal theory, can be seen as a special case of the X-curve suggested by reversal theory."
-
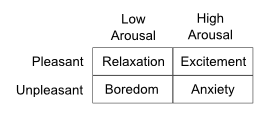 Figure 3a. (table) The very existence of colloquial labels for our emotions like relaxation, boredom, excitement and anxiety illustrates emotional/motivational bi-modality.
Figure 3a. (table) The very existence of colloquial labels for our emotions like relaxation, boredom, excitement and anxiety illustrates emotional/motivational bi-modality.

-
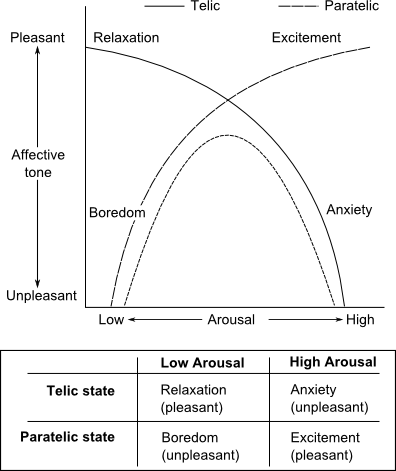 Figure 3a. (diagram with table) Each hypothetical curve represents the relationship between arousal and affective tone for one of the two stable states. These two states are labels "telic" (goal-directed; from the Greek word "telos" = "goal, end, close") and "paratelic" (behaviour-directed) respectively. (After Apter & Smith 1979, with permission).
Figure 3a. (diagram with table) Each hypothetical curve represents the relationship between arousal and affective tone for one of the two stable states. These two states are labels "telic" (goal-directed; from the Greek word "telos" = "goal, end, close") and "paratelic" (behaviour-directed) respectively. (After Apter & Smith 1979, with permission).

-
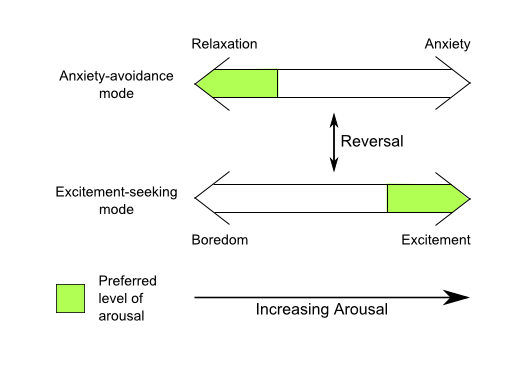 Figure 3b. At certain times the individual seeks high arousal which is then felt as pleasant when achieved ("excitement"); at other times he reverses to a state in which he seeks low arousal, at which time high arousal is felt as unpleasant ("anxiety"). In the former case low arousal is felt as unpleasant ("boredom") and in the latter case low arousal is felt as pleasant ("relaxation"). (After Apter & Smith, 1979).
Figure 3b. At certain times the individual seeks high arousal which is then felt as pleasant when achieved ("excitement"); at other times he reverses to a state in which he seeks low arousal, at which time high arousal is felt as unpleasant ("anxiety"). In the former case low arousal is felt as unpleasant ("boredom") and in the latter case low arousal is felt as pleasant ("relaxation"). (After Apter & Smith, 1979).

-
 Figure 3c. A commonly occurring Motivational Sequence.
Figure 3c. A commonly occurring Motivational Sequence.

".......... in the telic system the organism searches for security and safety in order to reduce arousal; in the paratelic system it searches for novelty and surprise in order to increase arousal. The latter is what is normally referred to as exploration. However, calling exploration a drive as it is now done widely, following the work of Butler (1953), Berlyne (1960), and others, implies that to discover new things is arousal reducing. Our claim is the contrary: namely that such discovery increases arousal and, in the paratelic mode, this is in fact pleasurable. This idea therefore constitutes an attack on the assumption that all behavior is governed by principles of drive-reduction (Smith & Apter, 1975).
The paratelic state is especially important for learning processes: "As defined earlier, the paratelic system is concerned with expressing behavior rather than goals and is perhaps seen in its purest state in children playing. It presumably arose because it had survival value in the young in allowing behavior to be exercised. In humans such play in the adult may be an example of neoteny which has turned out to be biologically advantageous"'' (Smith & Apter, 1975).
As in the theory of Behavioural Expansion, agonistic behaviors can be fitted very well into the theory of Psychological Reversals. Full- scale attack and flight are typically telic strategies in that they are of a strongly reactive nature, aimed at the achievement of a certain goal (subjugation or expulsion of a competitor, respectively escaping from a danger), and aimed at the reduction of the arousal level and re-stabil- ization of the status quo. Play-fight and social exploration are on the other hand typically paratelic strategies in that they are of a spontaneous non-reactive nature, aimed at raising the level of arousal.
4. An Additional Hypothesis: Integration of Theories on Aggression and on Learning
✰✰✰ <level 3> We postulate an additional hypothesis which allows us to integrate the foregoing theories more fully and which emphasizes the importance of stressful situations for learning.
In its simplest form this postulate states that in a stressful situation the individual arrives in a telic state and that in this telic (centripetal) state of emergency reactions, important neural and/or hormonal information is stored within the organism. This information is used automatically in the following non-stressful situation (paratelic state) to steer the growing process in the direction of optimal adaptation to the previously encountered stressful situation.
This process of the storage and subsequent processing and integration of information is carried through on an endocrine, neurophysiological and neural level or on either of these levels. Curiosity (paratelic) is steered in the direction of situations in which previously stress (with strong telic tendencies) has been experienced.
Expressed at the cognitive level it means that when this learning cycle runs optimally, very frightening situations gradually change into less frightening situations and eventually become intriguing. Having changed from frightening into intriguing, a situation attracts the individual's attention when he is in a paratelic (centrifugal) frame of mind. Thus additional experience with this particular situation is gathered until eventually the situation in question is not intriguing any more but evokes boredom when the individual is in a paratelic mood. At that stage the situation is of course not frightening any more either and the learning cycle has been completed. We could express this by saying that the stressful experience has been successfully 'digested', and the behavioural repertoire has accordingly 'expanded'.
Fig. 4 about here (Re-experiencing and proper digestion of an initially aversive experience; tabel/sequence)
(doc.121.pdf hier)
The hypothesis developed above implies that frightening and stressful situations can only be successfully 'digested' when they are followed by ample opportunity to arrive in a paratelic (centrifugal behavioural expansive state of mind).
A second prerequisite for a proper 'digestion' of experience is opportunity to tentatively reestablish contact with the source of previous trouble when in such a paratelic mood.
The adaptational value of such a system stems 'inter alia' from the fact that the learning process can be optimally fitted to varying circumstances and environments. When in a paratelic mood (surplus of energy available) the mechanism of preferential curiosity prevents the individual from wasting its energy and learning capacity on irrelevant objects and problems, but focuses the individual's attention on 'intriguing' environmental aspects which are, or have previously been, potential sources of trouble and stress, and therefore also potential sources of excitement.
The hypothesis also emphasizes the involuntary nature of learning: the individual willy-nilly launches itself periodically into trouble through paratelic action, and thus continues to accumulate experience in relevant areas of life.
5. Positive and Negative Learning Spirals
✰✰✰ <level 3> According to the presented hypothesis two learning spirals may be discerned, both of which result from positive feedback loops (see Fig. 4b and 4c). One spiral contains improper alternations of telic and paratelic
- Fig. 5a en 5b about here *********** Hier 2 figuren opnemen. De 1e is fig. 8 (Positive and negative learning spirals, showing alternations between telic and paratelic states)(doc.113.pdf)
De 2e figuur is fig.4 (Positive and Negative Learning Spirals) (Benoemen als fig.5a en fig.5b) (doc.116.pdf gebruiken; hier zitten een aantal slordigheden in; graag de teksten editen; die zijn nu vaak slecht leesbaar; ik stuur apart denk ik nog een betere scan (is gebeurd).)
states and results in ever more rigid and stereotyped behavior such as either immediate and full-scale flight or indiscriminate attack in agonistic situations, without options for intermediate responses and subtle alternations.
The other spiral contains proper alternations of telic and paratelic states and results in an ever growing diversity of adaptational responses (skills). The individual seeks to alternate the telic and paratelic states in an optimum rhythm and the higher his skills, the better he succeeds. Low skills on the other hand decrease the probability of maintaining a proper alternation of telic and paratelic states and thus enhance further attainment of rigid, stereotyped response-strategies instead of a growing diversity of optimal skills.
5.1 Genetical Aspects and 'Framing'
✰✰✰ <level 3> Whether an individual manages to deal with a situation successfully or not, depends - apart from learned skills - also on innate (constitutional, temperamental) qualities, like e.g. basic energy level, basic stability level, and specific learning dispositions. We may therefore expect that the rhythm of mood-reversals which is optimal for a certain type of experience differs from individual to individual right from the start. Furthermore, the 'critical frequencies' of experience may be expected to be different for every individual. And in the course of life these innate variations in 'critical frequencies' are further modified and/or amplified by processes of experience and learning.
Another way through which genetically determined predispositions come to life is through steering an individual's predilections whenever he is in a paratelic mood and seeks exciting experience. This process is sometimes referred to as 'framing' (Simpson, 1976).
As e.g. Waddington (1957) and Bateson (1976) point out when discussing views on behavioral development, it may be highly advantageous to combine concepts of predetermined, self-correcting features of developmental processes with concepts of adult behavior as the product of continuous interaction between the individual and its environment throughout ontogeny. As may be clear by now, the concept of learning as presented in the previous pages provides an excellent opportunity to reconcile such points of view that seemed incompatible in the heydays of the nature-nurture controversy.
5.2 Growth of Skills versus Growth of Unskills
✰✰✰ <level 3> Mastering of a certain setting generally implies the mastering of various sub-skills, some of which may be directly transferable as subroutines, required in other situations. Furthermore, acquired subroutines or sub-skills function as (emotional) refuge during the exploration of situations which require new abilities, but in which these subroutines may be applied. Through these two mechanisms, the transfer of subroutines and the improvement of the telic/paratelic balance, a high level of skills in a certain sub-domain enhances the growth of skills in related areas of life. Skills therefore will tend to grow in clusters.
But the growth of skills in other, quite different areas of life will be enhanced as well.
In short, the presented model explains on the one hand how situation-specific idiosyncracies in skills and unskills come about, and on the other hand predicts that individuals will tend to differ strongly in some overall level of skills. The latter, the general level of actualization of the behavioral repertoire, may, in our terms, also be formulated as the ability to effect a fluent, quick and agile shifting of the telic/paratelic (centrifugal/centrifpetal) equilibrium by able and subtle manipulation of the environment(ally induced aurousal), as immediate response to - even minute - changes in individual needs and motivational states.
The reverse side of the possibility of 'general actualization of individual abilities' is the possibility of general malfunctioning. The model predicts that malfunctioning in certain areas of life will enhance the malfunctioning in other areas of life as well. Lack of skills in one sub-setting drains the energy of an individual and evokes telic rather than paratelic behavioral strategies. This will result in less energy being available for investment in other settings and fewer paratelic moods which can result in exploring new options and solutions. Therefore skills enhance growth of further skills and 'unskills' enhance(2) further decrease of skills. Like skills, unskills also tend to grow in clusters, because faulty or deficient subroutines tend to influence the experience in related areas of life in a negative way. Such a cluster of 'unskills' might sometimes be labeled as 'phobia' or a specific form of neurosis, i.e., clusters of frightening and unmastered items, or specific memory constellations or, in the words of Grof (1972, 1973, 1976), negative 'COEX-systems' (systems of COndensed Experience). "The gradual successive growth of COEX systems by the mechanism of positive feedback (in the cybernetic sense) described, could account for the latency or 'incubation' period between the original traumatic events and future neurotic or even psychotic breakdowns. Manifest psychopatho- logical symptoms seem to occur at a time when the COEX system reaches a certain critical extension, and traumatic repetitions contaminate important areas of the patient's life and interfere with satisfaction of his basic needs" (Grof, 1976).
5.3 Reversability of 'Unskills'
✰✰✰ <level 3> As far as learning processes in Man are concerned, most 'unskills' are essentially reversible. Sometimes however, certain learning processes are constrained to specific stages of development. Much of the learning processes in Man is relatively free of this type of constraint (though it is not absent; see e.g. Benjamin, 1979), but in most animal species learning processes are very much dependent on the developmental stage (3) (age). One of the many experimental demonstrations of this principle has been provided by Kruijt (1964, 1971) who showed that certain behavioral unskills in social behavior of the jungle fowl cannot be cured by experience, once a certain stage of development has passed. His 'unskilled' cocks showed a type of social behavior which might very well be labeled as hopelessly neurotic. One of the conspicuous properties of their behavior was an exceptionally high frequency of violent aggression and fleeing during social encounters, at the expense of paratelic, appro¬priate (dis)play and courtship.
5.4 Displacement and Neuroses versus Psychological Health
✰✰✰ <level 3> Baerends (1976) discusses the phenomenon of 'displacement' behavior in animals and man. 'Displacement' is a label for functionally irrelevant behavior of an incomplete and ineffective character. He calls attention to the fact that such bursts of functionally irrelevant behavior in animals resemble neurotic symptoms in humans. Neuroses may, in this conception, be considered to be frequently reappearing sequences of stereotyped and irrelevant behavior. We might also describe them as negative COEX-systems or as sets of unskills.
Bloomfield et al.(1975) point out that in psychologically healthy individuals 'stay-and-play' responses (Orme-Johnson, 1973) are relatively more prominent than 'fight-or-flight' responses (Cannon, 1929). Psychological health implies so to speak the ability to endure the inevitable frustrations and irritations of everyday life without losing one's poise (Allport, 1961), and without being launched into a 'telic' mood. All-out aggression is therefore less likely to occur in 'self-actualizers' (Maslow, 1954) than in psychologically less healthy people.
6. The Use of the 'Aggression'-Label as an Element of the Agonistic Repertoire
✰✰✰ <level 3> Skills do not only influence the type of social role that an individual eventually manages to acquire, they are also of paramount importance for the flexibility with which the social roles in question are performed and adhered to. Psychological health and social skills enhance emancipated as well as emancipating behavior, and thus induce less rigid social role performances.
Benjamin's (1974, 1979) circumplex models of primate and human social behavior are based expl icitly on the notion that the style of performance in a dominant - as well as in a subordinate role depends strongly on the level of acquired (social) skills. The individual level of social skills (level of maturity) is, according to her one of the principal dimensions of social behavior. Social skills raise emancipation, acceptedness and autonomy, and diminish social conflict and role-interdependence .
Highly stereotyped and interdependent role-aspects such as scapegoat ing and mutual denunciation are therefore less likely to occur between highly (socially) skilled individuals than between psychologically less healthy people.
The use of the 'aggression'-lable may serve as an example. On the one hand a high-skilled individual will have to resort to agonistic actions less quickly if he wants to attain a certain social goal, because he will have a wider range of behavioral alternatives at his disposal. On the other hand, processes of mutual denunciation will be less obligatory in high-skilled individuals because of a lower interdependence. The tendency for stereotyped denunciation of the activities of competitors, superiors or critics as 'aggressive' will therefore be less strong, even in the case of strong coercive actions.
As far as the labels 'violent' and 'aggressive' are concerned, it is important to note that the behaviors which are indicated by these terms, can be described in positive terms as well. This can be achieved by simply considering the behavior in question from the point of view of a supporter. Any behavior, indicated as 'aggressive' or 'violent', appears to have reasonable components if regarded from a suitable viewpoint, and on the other hand no activity and no industry is, strictly speaking, possible without a certain amount of violence, no matter how little - as the archetypal pacifist Gandhi said.
The very subjectivity of the terms 'violence' and 'aggression' is highlighted by the complete chaos in aggression research, as far as the definitions of the basic concepts are concerned (see: van der Dennen, 1980).
Calling something 'aggression' implies some degree of derogation and is at variance with pure assessment of the behavior in question. Violence is mostly what others do to us, and aggression is very much in the eye of the beholder. The use of such depreciative, pejorative terms tells us therefore more about the nature of the user than about the behavior described. And in the case that 'violent' or 'aggressive' is used to describe a person, it gives us primarily information as to the (negative, dissociative) social relations between the rater and the ratee.
The use of these and similar depreciative and pejorative terms functions as a sort of involuntarily applied cognitive 'tool' on the level of social dynamics and interaction; a tool to create or maintain social distance on the physical or on the cognitive level.
This overt or internal-cognitive use of depreciatives establishes and consolidates social distance and alienation in the following way: The label 'aggressive' is used for example in some situation by A to denounce B's coercive actions. As soon as A has done that, he will as a consequence feel 'justified1 in seeking satisfaction for B's deeds. In that way A justifies also the coercive actions which he will subsequently put forth himself to harness, repel or constrain B. And A will probably not be inclined then to label his coercive action against B as 'aggressive'. For his own behavior he will tend, instead, to use a label which sounds more positive, such as 'tough' or 'energetic'. Thus a situation develops or is consolidated, in which each of the competitors considers himself as the 'good', and the other person as the 'bad' guy. And the net result is social distance and dissociation.
We could summarize these considerations by pointing out that 'aggression' in everyday parlance cannot be considered a proper and detached description of any behavior. Instead of serving the assessment of behavior, it serves the overruling of any detached appreciation of other persons' behavior by involuntary subjective distortion (4).
The use of the term 'aggression' may thus be a tool of the agonistic repertoire itself, serving to create or maintain distance, whether on the physical, the social or the cognitive level. Being an element of the agonistic repertoire, the use of the aggression-label often serves as a sort of emergency reaction on the level of social role behavior. The higher an individual's behavioral skill and mental health, the less he loses detachment and the less he tends to resort to (agonistic) emergency reactions, like using these labels. This is also acknowledged in the aggression theories of the so-called 'third trend' (Maslow, Fromm, Erikson, and others), an offshoot of self-actualization theory.
In this 'third trend', emphasis is laid on the negative correlation between an individual's tendency to apply (unnecessarily) violent actions and his level of psychological health and growth. The theory of aggression and learning processes as depicted in these pages may serve to support this axiom of the 'third-trend' investigations and in particular it may serve to explain why such a relationship between psychological growth and violent aggressiveness does exist.
7. By way of Conclusion
✰✰ <level 2> The reversal theory, applied to the notion of positive and negative learning spirals as presented in this paper, implies several predictions with respect to patterns of stereotyped 'agonistic' malfunctioning (5). The theory suggests that basic to any cure of an individual set of 'un-skills' is the recovery of a suitable telic/paratelic motivational balance. Only then may the individual establish proper alternations (see Fig. 3) of telic and paratelic states and thus profit from a growing variety of skills which are the result of positive learning spirals.
However, in order to recover a suitable telic/paratelic motivational balance, the individual must sufficiently often be able to attain relaxation of anxiety. And this is, as has been previously pointed out, exactly what is lacking in persons with a defective behavioral repertoire, whether or not labeled as 'aggressive' or 'violent'. The more an individual is in need of relaxation and reestablishment of a suit¬able telic/paratelic balance, the less he is therefore likely to succeed, and the more he will tend to resort to all-out reactive action to escape from sources of over-arousal. And social mechanisms of depreciation and repulsion may subsequently add to difficulties in obtaining relaxation. Whereas all-out reactive aggressive and violent action may appear quite necessary and justifiable to the actor in his 'telic' state, it may appear quite unfair, improper, inadequate or pathological to an onlooker.
This mechanism of the need for versus the availability of relaxation (Fig. 3), controlling the very basis of our behavioral growth, constitutes therefore an all-overruling source of systematic injustice. The better chances for learning are for those already favored. The evolutionary advantage of such a system of injustice may stem from the fact that it amplifies existing differences in quality and thereby intensifies existing selection pressures. Disadvantages on the genetic or cultural level will thus be eliminated more efficiently.
However advantageous this mechanism may be from an evolutionary point of view, it thoroughly thwarts any general effort to cure excessive amounts of aggressiveness. Annoying aggressiveness and agonistic 'over¬shooting1 may be viewed as outer manifestations of that amplified selection process. Maladaptive agressiveness evokes retaliation by others, or at least depreciation. And the very use of terms like 'aggressive' or 'pathological', by actors and reactors alike, enhances such depreciation and dissociation, thus making it even more difficult for the subject to regain social support, relaxation and subsequent paratelic states which are crucial for reestablishing positive learning spirals and the subsequent curing of maladaptive aggressiveness.
The common use in daily language of such labels is therefore not just a cognitive trap hampering a proper awareness of what is actually going on on the level of striving playing and learning, but a crucial link in the chain of social interactions which stabilizes social distance, negative learning spirals, and hampers the improvement of the very behavior which was labeled as aggressiveness or violence in the first place.
Hier een figuur 6 toevoegen. Daarvoor gebruiken: (doc.097.pdf en/of doc.098.pdf). Sommige onderdelen van de teksten zijn beter dan wel slechter leesbaar dan die in fig. 5 (doc.116.pdf). Bij het editen / verbeteren kan je teksten van elkaar "lenen" nof verwisselen naar smaak. Let er even op dat fig.5 en fig.6 t.o.v. elkaar de boven en onderkant verwisseld hebben.
Notes:
(1) This text has proceeded from a presentation at the 1978 congress of the "Ethologische Gesellschaft" at Basel, which presentation was partly supported by a grant from the "Fonds voor Nationale en Internationale Samenwerking" of the "Biologische Raad". It was presented at the First Congress of the European Section of the International Society for Research on Aggression, at Strassbourg, September 1981. The latter presentation was supported by a grant from the "Hendrik de Visser Fonds" of the Rijks Universiteit of Groningen to the senior author.
(2) 'Unskills' is defined here as learned (sub)routines with a high level of stereotypy and a low level of long-term efficiency and effectiveness. 'Unskills' are furthermore considered as fixated emergency-reactions.
(3) For well-known experiments with Rhesus monkeys, see e.g. Nowak & Harlow, 1975, or Suomi et al., 1976.
(4) To avoid confusion, one can rather resort to using the concept 'coercive power'. Tedeschi et al. (1974) have urged that a distinction be clearly drawn between the exercise of coercive power and the labeling of such action as aggressive by observers. According to these authors, coercion is a means of influence and when a person cannot persuade, bribe, manipulate, or otherwise induce a target to comply with his demands, then the success of influence may depend on the source's ability to restrain, transport, immobilize, injure or destroy the target. Coercion is often the influence strategy, in self-defense, for face-saving purposes, as an act of obedience to authority, in an attempt to reduce negative inequity, or for other reasons. "Perhaps the most important heuristic of adopting a language of coercive power is that it shifts attention from intrapsychic or biological determinants of behavior to the social causes. Instead of viewing harm doing as a manifestation of pent-up frustrations, Thanatos, or instinct, social psychological processes such as conflict, power, and normative demands for revenge or reciprocity become salient" (Tedeschi et al., 1977).
(5) See van der Dennen (1980) for a survey of the numerous categories of violent aggressiveness and related defective behaviour-patterns as established in current research on psychopathology and aggression.
References
ALLPORT, G.W. (1961) Pattern and Growth in Personality. Holt, Rinehart & Winston, N.Y.
APTER, M.J. & K.C.P. SMITH (1979) Sexual behaviour and the theory of psychological reversals. In: M. Cook & G. Wilson (Eds.) Love and Attraction. Pergamon, Oxford.
AZRIN, N.H., R.R. HUTCHINSON & R. McLAUGHLIN (1965) The opportunity for aggression as an operant reinforcer during aversive stimulation. Journal of the Experimental Analysis of Behavior 8: 171-180.
BAERENDS, G.P. (1960) Ethologie Compendium. Zool. Lab., Univ. Groningen.
BAERENDS, G.P. (1975) An evaluation of the conflict hypothesis as an explanatory principle for the evolution of displays. In: G. Baerends C. Beer & A. Manning (Eds.), Function and Evolution in Behaviour. Clarendon Press, Oxford.
BAERENDS, G.P. (1976) A tentative model for the causation of neuroses. In: H. van Praag (Ed.), Research in Neurosis. Bohn, Scheltema & Holkema, Utrecht.
BAERENDS, G.P., R. BROUWER & H.T. WATERBOLK (1955) Ethological studies on 'Lebistes reticulatus' (Peters) I. an analysis of the male courtship pattern. Behaviour, 8: 249-334.
BATESON, P.P.G. (1976) Rules and reciprocity in behavioral developments. In: P. Bateson & R. Hinde (Eds.) Growing Points in Ethology. Cambridge U.P.
BENJAMIN, L.S. (1974) Structural analysis of social behavior. Psychological Review, 81: 392-425.
BENJAMIN, L.S. (1979) Structural analysis of differentiation failure. Psychiatry, 42, 1: 1-23.
BERLYNE, D.E. (1960) Conflict, Arousal & Curiosity. McGraw-Hill, N.Y.
BERMOND, B. (1977) Horraonen en aggressief gedrag. In: P. Wiepkema & J. van Hooff (Eds.) Aggressief Gedrag; oorzaken en functies. Bohn, Scheltema & Holkema, Utrecht.
BLOOMFIELD, H.H., M.P. CAIN & D.J. JAFFE (1975) TM: discovering inner energy and overcoming stress. Dell Publ., N.Y.
BLURTON JONES, N.G. (1967) An ethological study of some aspects of social behaviour of children in nursery school. In: D. Morris (Ed.) Primate Ethology. Aldine, Chicago.
BLYTHE, P. (1973) Stress Disease. Baker, London.
BRANSON, F.H., M.H. STETSON & M.E. STIFF (1973) Serum FSH and LH in male mice following aggressive and nonaggressive interaction. Physiology & Behavior, 10, 2: 369-372.
BUCHHOLTZ, C.'(1978) Die Ethologie in der Lernforschung. Paper Ethologen-treffen, Basel.
BUSS, A.R. (1973) A conceptual framework for learning effecting the development of ability factors. Human development, 16: 273-292.
BUTLER, R.A. (1953) Discrimination learning by rhesus monkeys to visual exploration motivation. Journal of Comparative and Physiological Psychology, 46: 95-98.
CANNON, W.B. (1929) Bodily Changes in Pain, Hunger, Fear and Rage. Appleton, N.Y.
CHEREK, D.R., T. THOMPSON & G.T. HEISTAD (1973) Responding maintained by the opportunity to attack during an interval food reinforcement schedule. Journal of the Experimental Analysis of Behavior, 19: 113-123.
COLE, J.M. & B.K. PARKER (1971) Schedule-induced aggression: access to an attackable target bird as a positive reinforcer. Psychonomic Sciences, 22: 33-35.
CONNOR, J.L. (1974) Waning and recovery of conspecific aggression in the house mouse ('Mus musculus' L.). Journal of Comparative and Physiological Psychology, 87: 215-227.
CONNOR, J.L. & P. WATSON (1977) A device for presenting attack opportunity as a reinforcer for operant behavior. Behavioral Research Methods & Instrumentation, 9: 349-352.
CRAIG, W, (1918) Appetites and aversions as constituents of instincts. Biological Bulletin, 34: 91-107.
CRAIG, W. (1928) Why do animals fight? International Journal of Ethics, 31: 246-278.
CROOK, J.H. (1973) Darwinism and the sexual politics of primates. Social Science Information, 12: 7-28.
DAWKINS, R. (1976) The Selfish Gene. Oxford U.P., N.Y. van der DENNEN, J.M.G. (1980) Problems in the Concepts and Definitions of Aggression, Violence, and Some Related Terms. Polemological Institute, Univ. Groningen. van DIJK, J.J.M. (1977) Dominantiegedrag en Geweld: een multidisciplinaire visie op de veroorzaking van geweldmisdrijven. Dekker & van de Vegt, Nijmegen.
DREYER, P.I. & R.M. CHURCH (1970) Reinforcement of shock-induced fighting. Psychonomic Sciences, 18, 3: 14.7-148.
EIBL-EIBESFELDT, I. (1971) Ethology, the biology of behavior. Holt, Rinehart & Winston, N.Y.
EIBL-EIBESFELDT, I. (1975) Krieg und Frieden aus der Sichtder Verbaltens-forschung. Piper Verlag, München.
ELEFTHERIOU, B.E. & R.L. CHURCH (1968) Levels of hypothalamic luteinizing hormone-releasing factor after exposure to aggression (defeat) in C578BL/63 mice. Journal of Endocrinology, 42: 347-348.
FRY, D.P. (1980) The evolution of aggression and the level of selection controversy. Aggressive Behavior, 6, 1: 69-89.
GOODALL, J. (1971) Some aspects of aggressive behavior in a group of free-living chimpanzees. International Social Science Journal, 23, 1: 89-97.
GROF, S. (1972) Varieties of transpersonal experiences; observations from LSD psychotherapy. Journal of Transpersonal Psychology, 1972 (1).
GROF, S. (1973) Theoretical and empirical basis of transpersonal psychology and psychotherapy. Journal of Transpersonal Psychology, 1973 (1).
GROF, S. (1976) Realms of the Human Unconscious; observations from LSD research. Dutton, N.Y.
GROSS, H. (1978) Gewohnung beim Beutefang der Larve von Aeschua (Odonata) cyanea. Presentation at the 6th Ethologentreffen, 1978, at Basel.
HAMBURG, D.A. (1971) Recent research on hormonal factors relevant to human aggressiveness. International Social Science Journal, 23, 1: 36-47.
HAMILTON, W.D. (1971) Selection of selfish and altruistic behavior in some extreme models. In: J. Eisenberg & W. Dillon (Eds.) Man and Beast: comparative social behavior. Smithsonian Inst. Press, Washington D.C.
HARLOW, H.F. (1961) The development of affectional patterns in infant monkeys. In: B. Foss (Ed.) Determinants of Infant Behavior. Wiley, N.Y.
HARLOW, H.F. (1965) Total social isolation: effects on macaque monkey behavior. Science, 148: 666.
HINDE, R.A. (1960) Energy models of motivation. Symposia on Social Experimental Biology, 14: 199-213.
HINDE, R.A. (1966) Animal Behaviour: a synthesis of ethology and comparative psychology. McGraw-Hill, London (sec. ed. 1970).
HINDE, R.A. (1970) The nature and control of aggressive behavior. Paper UNESCO interdisciplinary expert meeting. Paris. (International Social Science Journal, 23(1), 1971).
HINDE, R.A. (197A) The study of aggression: determinants, consequences, goals and functions. In: J. de Wit & W. Hartup (Eds.) Determinants and Origins of Aggressive Behavior. Mouton, The Hague.
HOPF, S. (1981) Verhaltensbiologische Aspekte normaler und gestorter sozialer Entwicklung bei Primaten. In: Sozialisationsstorungen und Sucht. Akademische Verlagsgesellschaft, Wiesbaden.
KAVANAU, J. (1967) Behaviour of captive white-feeted mice. Science, 155: 1623-1639.
KELSY, J.E. & D. CASSIDY (1976) The reinforcing properties of aggressive vs. non-aggressive social interactions in isolated male ICR mice ('Mus musculus'). Aggressive Behavior, 2, A: 275-284.
KREUZ, L.E., R.M. ROSE & J.R. JENNINGS (1972) Suppression of plasma testosterone levels and psychological stress. Archives of General Psychiatry, 26: 479-482.
KRUIJT, J.P. (1964) Ontogeny of social behavior in Burmese red junglefowl ('Gallus gallus spadiceus') Bonnaterre. Behaviour, suppl. 12, whole volume.
KRUIJT, J.P. (1971) Ontogenie en gedrag, In: Ethologie, de biologie van gedrag. Pudoc, Wageningen.
KORTMULDER, K. (1972) A comparative study in colour patterns and behaviour in seven 'Barbus' species; a progress report. Behaviour, suppl. 19, whole volume.
KORTMULDER, K. (1974) On ethology and human behaviour. Acta Biotheoretica, 23, 2: 55-78.
LAGERSPETZ, K.M.J. (1964) Studies on the aggressive behavior of mice. Annales Academiae Scientiarum Fennicae, Series B, 131-133.
LEGRAND, R. (1970) Successful aggression as the reinforcer for runway behavior of mice. Psychonomic Sciences, 20, 5: 303-305.
LESHNER, A.I. (1975) A model for hormones and agonistic behavior. Physiology & Behavior, 15, 2: 225-235.
LEYHAUSEN, P. (1967) Zur Naturgeschichte der Angst. In: H. Wiesbrock (Ed.) Die Gesellschaftliche und Politische Rolle der Angst. Europäische Verlagsanstalt.
LORENZ, K. (1950) The comparative method in studying innate behaviour patterns. Symposia on Social Experimental Biology, 4: 221-268.
LORENZ, K. (1963) Das Sogenannte Böse; zur Naturgeschichte der Aggression. Borotha-Schoeck Verlag, Wien. MANNING, A. (1969) An Introduction to Animal Behaviour. Edward Arnold, London.
MARLER, P. & W.J. HAMILTON (1966) Mechanisms of Animal Behavior. Wiley, N.Y.
MARUYAMA, M. (1963) The second cybernetics: deviation-amplifying mutual causal processes. General Systems Yearbook, Vol. 8, 1963.
MASLOW, A.H. (1954) Motivations and Personality. Harper, N.Y. (-1970).
MASLOW, A.H. (1968) Toward a Psychology of Being. Van Nostrand, N.Y.
MAYNARD SMITH, J. (1974) The theory of games and the evolution of animal conflicts. Journal of Theoretical Biology, 47, 1: 209-221.
MAYNARD SMITH, J. (1978) The evolution of behavior. Scientific American, 239, 3: 136-145.
MAYNARD SMITH, J. & G.R. PRICE (1973) The logic of animal conflict. Nature, 246: 15-18.
van der MOLEN, P.P. (1979) Dimensions of social behavior in house mice ('Mus musculus'). Manuscript, Univ. Groningen. van der MOLEN, P.P. (1981) Striving, Playing and Learning: aggression and the dynamics of learning in the play and struggle called life. Manuscript, Univ. Groningen.
van der MOLEN, P.P. (in prep.) On the dimensions and dynamics of personality from mice to men. Heymans Bulletin, HB-78-401-EX, Univ. Groningen.
NOWAK, M.A. & H.F. HARLOW (1975) Social recovery of monkeys isolated for first year of life. I. Rehabilitation and therapy. Developmental Psychology, 11: 453-465.
0RME-JOHNSON, D.W. (1973) Autonomic stability and transcendental meditation. Psychosomatic Medicine, 35, 4: 341-349.
PARKER, G.A. (1974) Assessment strategy and the evolution of fighting behavior. Journal of Theoretical Biology, 47, 1: 223-243.
PEIJS, G.L.A.M. (1977) Development of Social Behaviour in the Rat. Nijmegen.
RASA, D.A.E. (1971) Appetence for aggression in juvenile Damsel fish. Zeitschrift fur Tierpsychologie, Beiheft 7, 1971.
RASA, D.A.E. (1980) Ethological aspects of aggressive behaviour. Paper Conference 'Biology of Aggression', Bonas, France, van ROOIJEN, J. (1976) 'Vlucht' en verwante begrippen in biologie en psychologie (Het Schade-anticiperend Gedragssysteem). Internal Report, Dept. of Ethology, Univ. Groningen.
ROSE, R.M., T.P. GORDON & I.S. BERNSTEIN (1972) Plasma testosterone levels in the male rhesus: influence of sexual and social stimuli. Science, 178: 643-645.
SCHUURMAN, T. (198]) Endocrine processes underlying victory and defeat in the male rat, Thesis, Univ. Groningen.
SCHUSTER, R.H. (1978) Ethological theories of aggression. In: I. Kutash et al. (Eds.) Violence: perspectives on murder and aggression. Jossey-Bass, San Francisco.
SCOTT, J.P. (1973) Hostility and Aggression. In: B. Wolman (Ed.) Handbook of General Psychology. Prentice-Hall, Englewood Cliffs.
SEVENSTER, P.A. (1973) Ethologie, de biologie van gedrag. Pudoc, Wageningen.
SIMPSON, M.J.A. (1976) The study of animal play. In: P. Bateson & R. Hinde (Eds.) Growing Points in Ethology. Cambridge U.P.
SMITH, K.C.P. & M.J. APTER (1975) A Theory of Psychological Reversals. Picton Publ., Chippenham.
SUOMI, S.J., M.L. COLLINS, H.F. HARLOW & G.C. RUPPENTHAL (1976) Effects of maternal and peer separations on young monkeys. Journal of Child Psychology and Psychiatry and Allied Disciplines, 17(2), 101-112.
TAYLOR, G.T. (1975) Male aggression in the presence of an estrous female. Journal of Comparative and Physiological Psychology, 89, 3: 246-252.
TEDESCHI, J.T., R.B. SMITH & R.C. BROWN (1974) A reinterpretation of re¬search on aggression. Psychological Bulletin, 81, 9: 540-562.
TEDESCHI, J.T., G.G. GAES & A.N. RIVERA (1977) Aggression and the use of coercive power. Journal of Social Issues, 33, 1: 101-125.
TELLEGEN, A. & J.M. HORN (1972) Primary aggressive motivation in three inbred strains of mice. Journal of Comparative and Physiological Psychology, 78, 2: 297-304.
TELLEGEN, A., J.M. HORN & R.G. LEGRAND (1969) Opportunity for aggression as reinforcer in mice. Psychonomic Sciences, 14: 104-105.
THOM, R, & E.C. ZEEMAN (1974) Catastrophe Theory: its present state and future perspectives. In: A. Manning (Ed.) Dynamical Systems. Springer Verlag.
THOMPSON, T.I. (1963) Visual reinforcement in Siamese fighting fish. Science, 141: 55-57.
THOMPSON, T.I. (1964) Visual reinforcement in fighting cocks. Journal of the Experimental Analysis of Behavior, 7: 45-49.
TINBERGEN, N. (1952) 'Derived' activities: their causation, biological significance, origin and emancipation during evolution. Quarterly Review of Biology, 27: 1-32.
TINBERGEN, N. (1954) The origin and evolution of courtship and threat displays. In: J. Huxley, A. Hardy & E. Ford (Eds.) Evolution as a Process. Allen & Unwin, London.
TINBERGEN, N. (1956) Fighting and threat in animals. New Biology, 14: 9-24.
TINBERGEN, N. (1969) The Study of Instinct. Oxford U.P., London. -
TRINCKER, D. (1966) Aufnahme, Speicherung und Verarbeitung von Information durch den Menschen. Veröffentlichungen der Schleswig-Holsteinischen Universitätsgesellschaft Neue Folge, 44.
WADDINGTON, C.H. (1957) The Strategy of the Genes. Allen & Unwin, London.
WIEPKEMA, P.R. & J.A.R.A.M. van HOOFF (Eds.) (1977) Agressief gedrag; oorzaken en functies. Utrecht, Bohn, Scheltema & Holkema.
WILSON, E.O. (1971) Competitive and aggressive behavior. In: J. Eisenberg & W. Dillon (Eds.). Man and Beast: comparative social behavior. Smithsonian Inst. Press, Washington D.C.
WILSON, E.O. (1975) Sociobiology; The New Synthesis. Harvard U.P., Cambridge, Mass.
ZEEMAN, E.C. (1976) Catastrophe Theory. Scientific American, 234, 4: 65-83.
ZILLMANN, D. (1979) Hostility and Aggression. Lawrence Erlbaum. Hillsdale N.J.